






SSE-V500
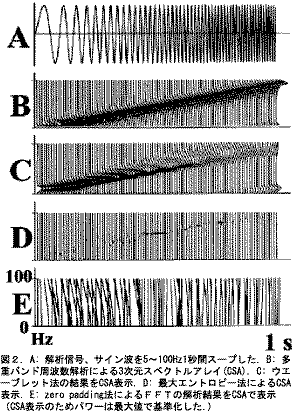
図2B-Eは解析信号を分析して3次元スペクトルアレイ(CSA)で表示した。図2B(MBFAによるCSA)に示しているように、10~100Hzまでのピークは直線的に増加したが10Hz以下のピークは非線形であった。図2Cでは10Hz以上の周波数におけるピークはほぼ直線的に増加したが、高い周波数では主要なパワーが減少し主要な周波数はステップ状に増加した。図2Dではいたるところでバラバラなピーク周波数によるパワーを示した。図2Eではパワーの正確な経緯を見ることができない。
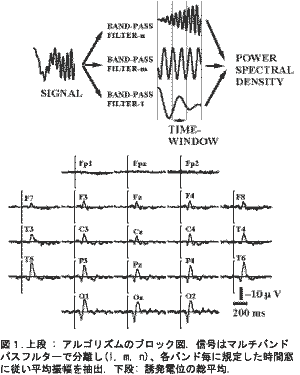
図1(下段)は6被験者総平均したVEPsです。110msの下向き陽性のピークと165msの上向き陰性のピークは右の後頭部(O2)で測定された、同様の応答は前額部(Fp1,Fpz,Fp2)以外の頭皮全体にわたって広くみられた。
図3(上段)は総平均したPSDsです;15Hz以上の帯域におけるすべてのサイトでパワーが非常に低く、15Hz以下の帯域はバックグラウンドパワーのみならず誘発パワーも高かった。誘発パワーはαおよびβ帯域において200ms付近で顕著であり、後頭部(O1,Oz,O2)で最大になった。
MANOVAは10~19Hz(df=3,103,p<0.05)において有意な結果を得た。1方向繰返し計測MANOVAはエポック間の著しい変化を示した(テーブル1)。刺激前と刺激後0-200msエポック間と、刺激後0-200msと刺激後200-400msエポック間において10Hz付近で有意な違いが得られた。18と19Hz付近では、刺激後200-400msと刺激後400-600ms間と、刺激前と刺激後0-200msエポック間において有意な変化が得られた。15Hz付近では、刺激後0-200msと刺激後200-400ms間と、刺激後200-400msと刺激後400-600msの間において有意な変化が得られた。被験者内有意差(df=3,60,p<0.05)を検定すると、エポックの影響は10-16,31-32,39-42,48-49,52-54,57-58,63-66,77-81,88-90Hzの帯域で有意であり、エポックとサイト間の相互作用は10-17,38-42,44,57-59,93Hzの帯域で有意であった。
図3(下段)は刺激前の平均値で基準化して総平均したPSDsである。αおよびβ帯域において200ms付近の誘発パワーは頭皮全体にわたって高まった、そして誘発パワーは前額部(F8,F4,Fz,F3,F7,Fp2,Fpz,Fp1)において有意であった。誘発パワーは100ms付近で生じ400ms付近まで継続した。
αおよびβ帯域における最大の誘発パワーは140ms付近の後頭部(O1,Oz,O2)に集中した。αおよびβ帯域におけるパワーの抑圧は後頭部において高い誘発パワーに続いて生じ、300msから記録終了(600ms)まで生じた。抑圧パワーは左右の後側頭部(T5,T6)で最大になった。40Hz付近の帯域パワーは後頭部(O1,Oz,O2)、頭頂部(P3,Pz,P4)、中央部(C3,Cz,C4)、前額部(F3,Fz,F4)において200ms以降に誘起され、後頭部(O1)において最大になった。30Hz以上の帯域パワーは中央部(C3,Cz,C4)、前額部(F3,Fz,F4)、頭頂部(P3,Pz,P4)において400ms以降に高くなった:このパワーは中央部で広く観察された。30Hz以上の帯域の誘発パワーは30~100Hzの帯域の間にとぎれとぎれに生じた。前額部(Fp1,Fpz,Fp2)のPSDsはどんな特徴的な変化も示さなかった。
考 察
多重バンド周波数解析
解析信号に関してMBFA法は1Hzと10ms分解能により良い結果のPSDsを得た;10Hz以上の帯域のピーク周波数は信号と線形な関係を示し、そのパワーは一定であった(図2B)。ウエーブレット解析[21]はパワーの変化が連続した時間に集中する傾向があり、周波数分解能は線形ではなく幾何級数的に変化する、そのため1Hzの分解能で信号を分析することは困難である。
テーブル1.統計解析の結果. サイト帯域(Hz)
*有意1:刺激前200msと刺激後0-200ms間の有意な変化
2:0-200msと200-400ms間の有意な変化
3:200-400msと400-600ms間の有意な変化
Burg法またはMEM[22,23]は滑らかな分解能で短い信号を解析する場合に使用する;しかしながら分析結果は使用するポールに非常に敏感でパワーは信号振幅を反映しない、したがってこれらの方法はパワーの時間経過を探求するには適切ではない。ZeroPadding法によるFFT[20]は短い時間窓に対応したものだが、PSDsを得ることは大変困難である(図2E)。

誘発関連電位PSDs
PSDsにおける絶対パワーは、15Hz以上の帯域と15Hz以下の帯域に明白な相違がある。 基準化したPSDs(図3下段)は絶対パワーPSDsと異なるパターンを示す;200ms付近の誘発パワーは前額部まで含む広い範囲で増加し、最大のパワーは後頭部で見られる。 誘発パワーに続く抑圧パワーはαおよびβ帯域で増加し、抑圧パワーは左右の後側頭部で最大になった。
以上2つの結果は後頭部による知見であるが、応答の分布は異なっている(テーブル1);エポック間(誘発前200msに対して誘発後0-200ms)の違いはP3,O1,O2,T5,T6上で有意である。 エポック間(誘発後200-400msに対して400-600ms)の違いはC3,C4,P3,P4,O1,O2,T5, T6,Pz,Oz上で有意であった。この結果は細胞集合体の2つの異なる機能を推測させた。 20Hz以上の帯域の誘発パワーは被験者間には有意ではないが、被験者内には有意である。γ帯域の正確な誘発の変動は被験者間に大きく、時間的経過も複雑である。
γ帯域の範囲
JasperとAndrews[25]はγリズムが30か35Hz以上の周波数に存在すると報告した。しかし最近はγ周波数が単にβ帯域の一部であるとみなしている[26]。γ帯域の範囲の報告は様々である; 20-50 [6],30-70 [27], 30-110 [15],40Hz以上[28]。 Crone達[13]によれば、γ帯域は皮質の形成状態から2つの帯域に分類できることを報告した、γ帯域は脳機能の研究により詳細に分類しなければならない。 FFTにおいて周波数分解能は時間窓に制限され、時間分解能を狭めに選択すると周波数分解能が広がる。したがってγ帯域の状態をFFTにより解析することはできない。 しかしMBFAを用いると1Hzと10ms分解能でPSDsが得られ、しかも1Hzと10msの2つのパラメータは互いに依存しない。
同期と非同期
γ帯域の応答に関する報告は帯域の同期性について論じているが、いくつかの研究は脳機能における非同期性と関係付けて論じている[9,13]。ここで誘発の高いパワーは同期と対応し、非同期は抑圧パワーと対応すると考えられる。1Hzと10ms分解能によるPSDsは複雑なパターンを示す:特定の帯域のパワーは高く、いくつかの帯域は変化がなく、その他の帯域は抑圧されていた。
γ帯域の分布
γ帯域の分布は瀰漫性に広く観察されると報告されている[29,30]。 MBFAによるとγ帯域の同期では広く瀰漫性に分布している、γ帯域の同期とαおよびβ帯域とは異なった分布を示す。
γ帯域の時間
γ帯域の変動は、猫の聴覚システムにおいて刺激後の100msから生じると報告がある [29]、しかし名詞と動詞による人間のγ帯域の変動は刺激後500-800msに生じると報告されている[7]。MBFAは200ms以降の40Hz付近の帯域におけるγ帯域の同期と、400ms以降のその他のγ帯域の同期を明らかにした。
課題とγ帯域
人間の視覚機能において、30-40HzEEG活動は短時間のパターン刺激により誘発されることが報告されている[3]。猫の大脳皮質において、細胞間の情報のやりとりが、γ帯域のゆらぎを発生させている[31]。 γ帯域の変動は異なった細胞集合体から発生し、緊張した主要な部位の状態[10,28]、言語認識[7,32]、特定言語プロセス[33]、複数パターンの認知[34]、知覚と記憶の構造、言語的プロセス、その他行動状態と知覚機能[35]に依存する。ほとんどの著者がγ帯域の応答[27,36]は認識や知覚の高度な脳機能によると報告しているが、PSDsはγ帯域がフラッシュ注視という簡単な課題に対してさえ複雑に応答することを詳細に示している。
誘発電位とγ帯域
誘発電位の発生源(N1)とγ帯域の変動は異なる[37]。イベントに関連した電位は200ms付近で活性化する、そして刺激後500-800msにおける30Hz付近のγ帯域の変動は連続した活動と考えられ、細胞集合体の2つの異なる機能的な状態を示す[7]。 MBFAによれば15Hz以下の帯域(100-400ms)における誘発パワーはVEPs(P110とN165)の顕著なピークに一致する、そして15Hz以下の誘発パワーの分布はVEPsのピークに一致する。しかし30Hz以上のγ帯域はVEPsのピークとは異なった分布を示す;γ帯域の同期は200ms以降に生じ、VEPsのピークとは異なる分布を示す。
αおよびβ帯域に対するγ帯域
γ帯域の活性化はα帯域の変化から独立していると報告されている[14]、イベントに関連して同期した低いγ帯域と、イベントに関連した非同期のα帯域は平行して観察されている[13]。誘発の高いパワーと抑圧されたパワーは異なる分布になる、基準化されたPSDsではαおよびβ帯域において異なる時間的な経過を示す。しかしγ帯域の同期と、αおよびβ帯域の非同期は同じ分布にはならない。さらに我々はγ帯域の同期とαおよびβ帯域の非同期の間では異なる分布であることを導きだした。これは異なる細胞集合体が引き起したことを示唆している。
結 論
結論として、フラッシュのような簡単な刺激においてさえMBFAによって解析された詳細なPSDsは、αおよびβ帯域の同期と、異なるエリア上で2つの応答による非同期が生じることが知られた。 γ帯域の同期は詳細に観測され、様々な帯域が対象によって引き起された。 MBFA法はγ帯域のみならずαおよびβ帯域においても、同期と非同期を詳細に導きだした。
マルチバンドパスフィルターを用いた
フラッシュ誘発電位の高精度スペクトル解析
![]()
要 約
高速フーリエ変換(FFT)はパワースペクトルの分布(PSD)を知るには良い方法だが周波数分解能がサンプリング窓により制限される。短時間の信号では詳細な周波数情報は得られない。MBFA(MultiBandpassFilterAnalysis)法を用いることによりフラッシュによる視覚誘発電位(VEPs)からFFTの限界を超えて詳細なPSDが得られた。脳波は平衡型頭部外基準で国際10-20法により21部位から記録し、-200~600ms間を1,000Hz12bitsでサンプルした。PSDは10~100Hzを周波数分解能1Hz、時間分解能10msで解析した。αとβ帯域の背景波は後頭部で強く認められたが、刺激後200ms付近に、VEPsのP110とN165に相当すると考えられるパワーのさらなる増加が後頭部で観察された。刺激前の平均値で基準化したPSDでは、後頭部で200ms付近の誘発反応と、それに続いた抑制反応が強調された。20Hz以上の帯域は統計上有意ではないが、γ帯域は同じ刺激でも誘発周波数が個人内で毎回変動し、6被験者の個人差も認められた。PSDの細かな変化はフラッシュ注視という単純な課題でさえ複雑であり、MBFAにより時間的経過と詳細なスペクトル分布が観察された。
![]()
γ帯域の変動は脳組織[1]や、猿[2]・人間[3-8]で研究されているが、γ帯域における変動の詳細は明確ではない。多くの論文は脳の活性とγ帯域の同期について論じているが、いくつかのグループは課題の差による非同期性を報告している[9,10]。γ帯域の変動する時間は課題により異なっており[11]、異なる周波数が異なった領域上で発生すると報告している[12]。γ帯域の時間と空間上の位置は、高低のγ帯域により同期が異なり、低いγ帯域の同期はα帯域の非同期とパラレルである[13]。γ帯域の変動とα帯域の変動は2つの独立した脳の活動と、2つの異なった機能に影響を受けている[14]。γ帯域活性の周波数は知覚の状態に依存することが知られている[15]。γ帯域の分析では高速フーリエ変換(FFT)によりパワースペクトル(PSD)を得る方法は有用であるが[16,17]、FFTは周波数分解能が時間分解能と背反するため限界がある[周波数分解能]=1/[時間分解能]。時間と周波数の分解能は時系列信号の分析にとって重要である。そのためFFTは信号の詳細な変化を知るには適切な方法ではない[18]。スペクトルの時間的な変化はランニングスペクトル[8,13,19]とZeroPadding[20]法により得ている。しかしこの方法は、解析時間幅(FFTのポイント数)が時間分解能より広い場合は同じデータを繰り返し使用するため結果が平均され低SN比をもたらす。ウエーブレット[21]と最大エントロピー法(MEM)[22,23]は短い信号を解析するのに用いられている。これらの方法は複数のパラメータの影響を大きく受け、パワーの時間的な経過を知るには不適切である。FFTに依存しない新しいアルゴリズムは、FFTの限界を超えてVEPsの詳細な時間経過を知るのに有用である。MBFA法はフラッシュ注視という簡単な課題において1Hzの分解能でPSDsが得られるよう導入した。そしてVEPsにおける時間と空間上の情報が得られた。
対象と方法
対象:6人のボランティア33.8±9.5歳(mean±SD)が、全ての研究に対する完全な説明の後に自発的に同意してこの研究に参加した。視力は正常であるか適切に度数を補正した。
課題:30分間の暗順応した後、対象から1mの位置に設置したストロボスコープ(0.1J,3×3cm)を両目で見るように指示した。刺激間隔は2~4秒間で、1対象に50回以上の刺激を与え、アーチファクトを除去するよう慎重に観察した後に50回以上の誘発脳波を検討した。
記録:平衡型頭部外基準で国際10/20システムに順じた21点頭皮電極を用いた[24]。信号は0.5~100Hz(-6dB/octave)を1,000Hzサンプリングして、刺激前200ms刺激後600ms間記録した。ゼロレベルは各信号の刺激前200msの平均電位を用いた。
アルゴリズム:多重バンド周波数解析(MBFA):図1(上段)はこのアルゴリズムのブロック図で、バンドパスフィルタにより信号を処理し、全ての帯域において指定した時間幅の平均振幅を求める。抽出する時間幅が波長の幅より短い場合は、波形の山と谷から平均振幅を求める。1,000Hzサンプリングにおける最小の周波数分解能は1Hzで、最も低い帯域は4.5-5.5Hz、最も高い帯域は299.5-300.5Hz、最小のスペクトル抽出時間幅は10msである。
利点:MBFAは同じ周波数分解能においてFFTの50倍の時間分解能が得られ、結果はFFTと同じフーリエ級数のスペクトルである。全ての周波数において解析における過渡特性が同一である、これは周波数にかかわらず時間情報が正確に得られることを意味し、周波数の時間的変動や周波数間の発生時間差が詳細に把握できる。時間窓を周波数分解能と無関係に設定でき、解析対象や目的に合せて特性を変えられる。限られた区間を解析対象としないので、窓関数やアンチエリアシングフィルターが不要で歪が生じない。低い周波数の半波長であっても時間的な拡散が生じず正しい振幅と時間の情報が得られる。
欠点:MBFAは解析結果に位相情報を持っていない、このため逆変換ができない。
手順:解析信号は正弦波を5~100Hz1秒間掃引した波形(マルチファンクションシンセサイザー1941、NF回路ブロック(株)、日本)を、12ビット1,000Hzでサンプリングして作成した(図2A)。解析信号は、MBFA法、ウエーブレット法、最大エントロピー法(MEM)、ZeroPadding法によるFFTを使用して分析した。VEPsにおいて平均したPSDsを得るために50回の誘発電位をMBFA法で解析した。基準化したPSDsは、各帯域に対応する刺激前200msの平均パワーを基準にしてデシベルに変換した。各10msの信号はデータのオーバーラップ無しに、1Hz10ms分解能で連続して解析した。全体を平均したVEPsはPSDsと比較して計算した。
統計:分散の多変量解析は(MANOVA)、200ms間の平均振幅をエポックとして各帯域が記録した変化をサイトと検定した(4エポック×21サイト、Systat5.2.1)。有意な帯域においては、1方向繰返し計測MANOVAを使用し、各サイトを用いてエポックの有意性を評価した(4エポック)。分散分析(ANOVA)は被験者内の変化を評価するのに用い、主要な同期した誘発パワーにおける各帯域の効果を検定した(4エポック×21サイト、Fisher'sLSD)。
結 果
 |
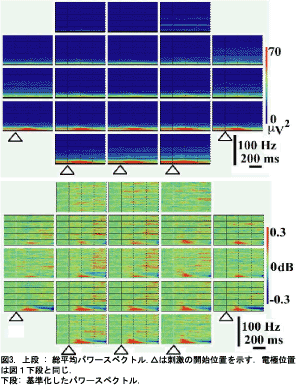 |
図2B-Eは解析信号を分析して3次元スペクトルアレイ(CSA)で表示した。図2B(MBFAによるCSA)に示しているように、10~100Hzまでのピークは直線的に増加したが10Hz以下のピークは非線形であった。図2Cでは10Hz以上の周波数におけるピークはほぼ直線的に増加したが、高い周波数では主要なパワーが減少し主要な周波数はステップ状に増加した。図2Dではいたるところでバラバラなピーク周波数によるパワーを示した。図2Eではパワーの正確な経緯を見ることができない。
図1(下段)は6被験者総平均したVEPsです。110msの下向き陽性のピークと165msの上向き陰性のピークは右の後頭部(O2)で測定された、同様の応答は前額部(Fp1,Fpz,Fp2)以外の頭皮全体にわたって広くみられた。
図3(上段)は総平均したPSDsです;15Hz以上の帯域におけるすべてのサイトでパワーが非常に低く、15Hz以下の帯域はバックグラウンドパワーのみならず誘発パワーも高かった。誘発パワーはαおよびβ帯域において200ms付近で顕著であり、後頭部(O1,Oz,O2)で最大になった。
MANOVAは10~19Hz(df=3,103,p<0.05)において有意な結果を得た。1方向繰返し計測MANOVAはエポック間の著しい変化を示した(テーブル1)。刺激前と刺激後0-200msエポック間と、刺激後0-200msと刺激後200-400msエポック間において10Hz付近で有意な違いが得られた。18と19Hz付近では、刺激後200-400msと刺激後400-600ms間と、刺激前と刺激後0-200msエポック間において有意な変化が得られた。15Hz付近では、刺激後0-200msと刺激後200-400ms間と、刺激後200-400msと刺激後400-600msの間において有意な変化が得られた。被験者内有意差(df=3,60,p<0.05)を検定すると、エポックの影響は10-16,31-32,39-42,48-49,52-54,57-58,63-66,77-81,88-90Hzの帯域で有意であり、エポックとサイト間の相互作用は10-17,38-42,44,57-59,93Hzの帯域で有意であった。
図3(下段)は刺激前の平均値で基準化して総平均したPSDsである。αおよびβ帯域において200ms付近の誘発パワーは頭皮全体にわたって高まった、そして誘発パワーは前額部(F8,F4,Fz,F3,F7,Fp2,Fpz,Fp1)において有意であった。誘発パワーは100ms付近で生じ400ms付近まで継続した。
αおよびβ帯域における最大の誘発パワーは140ms付近の後頭部(O1,Oz,O2)に集中した。αおよびβ帯域におけるパワーの抑圧は後頭部において高い誘発パワーに続いて生じ、300msから記録終了(600ms)まで生じた。抑圧パワーは左右の後側頭部(T5,T6)で最大になった。40Hz付近の帯域パワーは後頭部(O1,Oz,O2)、頭頂部(P3,Pz,P4)、中央部(C3,Cz,C4)、前額部(F3,Fz,F4)において200ms以降に誘起され、後頭部(O1)において最大になった。30Hz以上の帯域パワーは中央部(C3,Cz,C4)、前額部(F3,Fz,F4)、頭頂部(P3,Pz,P4)において400ms以降に高くなった:このパワーは中央部で広く観察された。30Hz以上の帯域の誘発パワーは30~100Hzの帯域の間にとぎれとぎれに生じた。前額部(Fp1,Fpz,Fp2)のPSDsはどんな特徴的な変化も示さなかった。
考 察
多重バンド周波数解析
解析信号に関してMBFA法は1Hzと10ms分解能により良い結果のPSDsを得た;10Hz以上の帯域のピーク周波数は信号と線形な関係を示し、そのパワーは一定であった(図2B)。ウエーブレット解析[21]はパワーの変化が連続した時間に集中する傾向があり、周波数分解能は線形ではなく幾何級数的に変化する、そのため1Hzの分解能で信号を分析することは困難である。
テーブル1.統計解析の結果. サイト帯域(Hz)
| 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | |
|---|---|---|---|---|---|---|---|---|---|---|
| FP1 | ||||||||||
| Fp2 | ||||||||||
| F3 | ||||||||||
| F4 | *(2) | *(2) | * | |||||||
| C3 | *(3) | *(3) | ||||||||
| C4 | * | *(3) | *(3) | |||||||
| P3 | *(1,2) | *(1) | * | *(1,3) | *(1,3) | |||||
| P4 | *(3) | |||||||||
| 01 | *(1) | *(3) | *(3) | |||||||
| 02 | *(1,2) | *(1,2,3) | *(2,3) | *(1,3) | *3 | |||||
| F7 | ||||||||||
| F8 | ||||||||||
| T3 | ||||||||||
| T4 | ||||||||||
| T5 | *(1,2) | *(1,2) | *(1,3) | *(1,3) | *(1,3) | |||||
| T6 | *(1) | *(3) | *(3) | |||||||
| Fpz | ||||||||||
| Fz | ||||||||||
| Cz | ||||||||||
| Pz | *(3) | |||||||||
| Oz | *(3) |
*有意1:刺激前200msと刺激後0-200ms間の有意な変化
2:0-200msと200-400ms間の有意な変化
3:200-400msと400-600ms間の有意な変化
Burg法またはMEM[22,23]は滑らかな分解能で短い信号を解析する場合に使用する;しかしながら分析結果は使用するポールに非常に敏感でパワーは信号振幅を反映しない、したがってこれらの方法はパワーの時間経過を探求するには適切ではない。ZeroPadding法によるFFT[20]は短い時間窓に対応したものだが、PSDsを得ることは大変困難である(図2E)。
誘発関連電位PSDs
PSDsにおける絶対パワーは、15Hz以上の帯域と15Hz以下の帯域に明白な相違がある。 基準化したPSDs(図3下段)は絶対パワーPSDsと異なるパターンを示す;200ms付近の誘発パワーは前額部まで含む広い範囲で増加し、最大のパワーは後頭部で見られる。 誘発パワーに続く抑圧パワーはαおよびβ帯域で増加し、抑圧パワーは左右の後側頭部で最大になった。
以上2つの結果は後頭部による知見であるが、応答の分布は異なっている(テーブル1);エポック間(誘発前200msに対して誘発後0-200ms)の違いはP3,O1,O2,T5,T6上で有意である。 エポック間(誘発後200-400msに対して400-600ms)の違いはC3,C4,P3,P4,O1,O2,T5, T6,Pz,Oz上で有意であった。この結果は細胞集合体の2つの異なる機能を推測させた。 20Hz以上の帯域の誘発パワーは被験者間には有意ではないが、被験者内には有意である。γ帯域の正確な誘発の変動は被験者間に大きく、時間的経過も複雑である。
γ帯域の範囲
JasperとAndrews[25]はγリズムが30か35Hz以上の周波数に存在すると報告した。しかし最近はγ周波数が単にβ帯域の一部であるとみなしている[26]。γ帯域の範囲の報告は様々である; 20-50 [6],30-70 [27], 30-110 [15],40Hz以上[28]。 Crone達[13]によれば、γ帯域は皮質の形成状態から2つの帯域に分類できることを報告した、γ帯域は脳機能の研究により詳細に分類しなければならない。 FFTにおいて周波数分解能は時間窓に制限され、時間分解能を狭めに選択すると周波数分解能が広がる。したがってγ帯域の状態をFFTにより解析することはできない。 しかしMBFAを用いると1Hzと10ms分解能でPSDsが得られ、しかも1Hzと10msの2つのパラメータは互いに依存しない。
同期と非同期
γ帯域の応答に関する報告は帯域の同期性について論じているが、いくつかの研究は脳機能における非同期性と関係付けて論じている[9,13]。ここで誘発の高いパワーは同期と対応し、非同期は抑圧パワーと対応すると考えられる。1Hzと10ms分解能によるPSDsは複雑なパターンを示す:特定の帯域のパワーは高く、いくつかの帯域は変化がなく、その他の帯域は抑圧されていた。
γ帯域の分布
γ帯域の分布は瀰漫性に広く観察されると報告されている[29,30]。 MBFAによるとγ帯域の同期では広く瀰漫性に分布している、γ帯域の同期とαおよびβ帯域とは異なった分布を示す。
γ帯域の時間
γ帯域の変動は、猫の聴覚システムにおいて刺激後の100msから生じると報告がある [29]、しかし名詞と動詞による人間のγ帯域の変動は刺激後500-800msに生じると報告されている[7]。MBFAは200ms以降の40Hz付近の帯域におけるγ帯域の同期と、400ms以降のその他のγ帯域の同期を明らかにした。
課題とγ帯域
人間の視覚機能において、30-40HzEEG活動は短時間のパターン刺激により誘発されることが報告されている[3]。猫の大脳皮質において、細胞間の情報のやりとりが、γ帯域のゆらぎを発生させている[31]。 γ帯域の変動は異なった細胞集合体から発生し、緊張した主要な部位の状態[10,28]、言語認識[7,32]、特定言語プロセス[33]、複数パターンの認知[34]、知覚と記憶の構造、言語的プロセス、その他行動状態と知覚機能[35]に依存する。ほとんどの著者がγ帯域の応答[27,36]は認識や知覚の高度な脳機能によると報告しているが、PSDsはγ帯域がフラッシュ注視という簡単な課題に対してさえ複雑に応答することを詳細に示している。
誘発電位とγ帯域
誘発電位の発生源(N1)とγ帯域の変動は異なる[37]。イベントに関連した電位は200ms付近で活性化する、そして刺激後500-800msにおける30Hz付近のγ帯域の変動は連続した活動と考えられ、細胞集合体の2つの異なる機能的な状態を示す[7]。 MBFAによれば15Hz以下の帯域(100-400ms)における誘発パワーはVEPs(P110とN165)の顕著なピークに一致する、そして15Hz以下の誘発パワーの分布はVEPsのピークに一致する。しかし30Hz以上のγ帯域はVEPsのピークとは異なった分布を示す;γ帯域の同期は200ms以降に生じ、VEPsのピークとは異なる分布を示す。
αおよびβ帯域に対するγ帯域
γ帯域の活性化はα帯域の変化から独立していると報告されている[14]、イベントに関連して同期した低いγ帯域と、イベントに関連した非同期のα帯域は平行して観察されている[13]。誘発の高いパワーと抑圧されたパワーは異なる分布になる、基準化されたPSDsではαおよびβ帯域において異なる時間的な経過を示す。しかしγ帯域の同期と、αおよびβ帯域の非同期は同じ分布にはならない。さらに我々はγ帯域の同期とαおよびβ帯域の非同期の間では異なる分布であることを導きだした。これは異なる細胞集合体が引き起したことを示唆している。
結 論
結論として、フラッシュのような簡単な刺激においてさえMBFAによって解析された詳細なPSDsは、αおよびβ帯域の同期と、異なるエリア上で2つの応答による非同期が生じることが知られた。 γ帯域の同期は詳細に観測され、様々な帯域が対象によって引き起された。 MBFA法はγ帯域のみならずαおよびβ帯域においても、同期と非同期を詳細に導きだした。